О происхождении народов и ДНК-генеалогии

Отредактированный фрагмент из книги Олега Валецкого и Браниславы Совиль «Сербская руна», вышедшей на сербском языке в Белграде в 2014 году в издательстве «Пешич и синови».
Ныне ДНК-генеалогия является уже признанным направлением науки, наряду с ДНК-тестированием (которое, строго говоря, не является ДНК-генеалогией, и направлено на получение гаплотипов и гаплогрупп, прежде всего в США, где оно и получила самое большое развитие[1]). По сути, ДНК-генеалогия — историческая наука, как подробно разъяснено ниже.
В России ДНК генеалогия стала получать популярность благодаря профессору Анатолию Клёсову, переехавшему на работу в США еще во времена СССР как специалист-биохимик по межгосударственному соглашению. В 2015 году он основал в России Академию ДНК-генеалогию и создал Лабораторию ДНК-генеалогии, где проводится массовое тестирование добровольцев и проводится персональная интерпретации и историческая реконструкция получаемых данных.
В США, где Клёсов, профессор Гарварда, разрабатывал лекарства против рака, а также создавал новые композиционные материалы. ДНК-генеалогией он занялся уже после того, как получил признание данных областях в США. В России он стал известен как раз благодаря ДНК-генеалогии. В СССР он был удостоен звания лауреата Государственной премии СССР как биохимик.
Анатолий Клёсов успешно продолжает вести научно-исследовательскую работу, главным образом на собственные средства, зарабатываемые им в США.
На международной конференции в Сербии «У источника культуры и науки», прошедшей в Белграде с 21 по 23 сентября 2012 года[2] Клёсов представил доклад «ДНК-генеалогия предков славян со времен Лепенского Вира и Винчи».
В докладе были представлены данные о древности арийского эпоса на основании астрономических данных, упоминавшихся в индийских ведах, показаны гаплогруппы и гаплотипы жителей Балкан и Сербии, в частности, приведено дерево 185 гаплотипов Сербской Воеводины. По результатам конференции Анатолий Клёсов опубликовал в издаваемом им журнале «Вестник Академии ДНК-генеалогии»[3], выходящем в Бостоне и в Москве статью «Конференция в Белграде по истокам культуры и науки (21–23 сентября 2012) на фоне общих проблем в Сербии».
Вот что пишет сам Анатолий Клёсов о том, что такое ДНК-генеалогия:
«ДНК-генеалогия, или, условно говоря, “молекулярная история”, оперирует объективными данными. Они формируются в виде мутаций в ДНК, что дает возможность в свою очередь оперировать гаплогруппами — родами человеческими, и гаплотипами — индивидуальными характеристиками ДНК. То есть ДНК-генеалогия дает возможность оперировать объективными генеалогическими “паспортами”, которые тысячелетиями и десятками тысяч лет передаются по наследственной цепочке и позволяют видеть древнюю историю, древние миграции в пространстве и во времени. Такой подход дает принципиально новую возможность открыть, описать, рассмотреть историческую проблему под новым, необычным углом.
На чем же основывается ДНК-генеалогия? Экспериментальные данные ДНК-генеалогии — это картина мутаций в мужской половой хромосоме, или Y-хромосоме. Эту картину мутаций ДНК-генеалогия рассматривает как в Y-хромосомах отдельных людей, так и их групп, популяций. Методология новой науки — перевод динамической картины мутаций в хронологические показатели, во времена жизни общих предков популяций, а значит — общих предков древних родов и племен. То есть фактически производится расчет времен, когда в древности жили эти рода и племена. В свою очередь, построение “карты” этих времен по регионам, материкам, континентам позволяет понять миграционные пути наших предков.
Мутации, рассматриваемые в ДНК-генеалогии — это или одиночные (как правило) замены нуклеотидов в ДНК, как, например, аденин на цитозин, или цитозин на тимин, или вставки нуклеотидов, или делеции, или мутации более сложные, при которых ошибка копирующего фермента приводит к переносу целого блока нуклеотидов, тандемного, как его порой называют. Это дает или удлинение серии таких блоков на один (редко — сразу на два-три блока), или их укорачивание. Эти мутации наследуются в поколениях, порой в сотнях и тысячах поколений подряд без изменений, или с небольшими вариациями, и тем самым служат надежными, порой “вечными” метками в ДНК.
Первые мутации, или “снипы”, обычно очень стабильны, и их для ДНК-генеалогии отбирают только такие, которые случаются только один раз (два раза — максимум) за историю человечества. Поэтому они являются маркерами родов человечества. Эти рода называют “гаплогруппами”, каждая из которых состоит из многих выявленных подгрупп, но все подгруппы несут родо-определяющую (или гаплогруппу-определяющую) мутацию. Всего в мире насчитывают 20 основных родов, обозначаемых буквами латинского алфавита, от А до Т.
Мутации второго типа значительно более быстрые, и происходят в определенных локусах ДНК (то есть в гаплотипах, которые и есть совокупность выбранных локусов) раз в несколько десятков или сотен поколений. Поэтому гаплотипы ДНК выбирают так, чтобы в них таких локусов было как можно больше, и сейчас работа рутинно ведется с 67-маркерными гаплотипами (правда, в академических публикациях это обычно от 8 до 17 маркерных гаплотипов), а последние два года — и с 111-маркерными гаплотипами. В 67-маркерных гаплотипах, например, одна мутация происходит в среднем за 8 поколений, в 111-маркерных — за 4 поколения, то есть примерно раз в сто лет.
Поскольку уже показано и доказано, что эти мутации в локусах происходят в основном (или исключительно) неупорядоченно, то к ним оказалось возможным применять правила и подходы химической кинетики (или биологической кинетики, которая базируется на тех же принципах химической кинетики, только в биологических объектах). Каждому маркеру свойственна определенная константа скорости мутации (измеряемая в числе мутаций на маркер на 25 лет, то есть на условное поколение), и каждому гаплотипу (состоящему из набора маркеров) — тоже свойственна определенная средняя константа скорости мутации. Поэтому чем древнее общий предок популяции, тем больше мутаций накапливается в гаплотипах его потомков по сравнению с ним, тем больше поколений (и лет) отделяет современных потомков от их общего предка, и это число поколений (и лет) рассчитывается методами ДНК-генеалогии.
В классификации ДНК-генеалогии рода, они же гаплогруппы, различаются по мутациям: все без исключения “восточные славяне” (гаплогруппа R1a у славян) имеют снип М198, все “северо-западные славяне” (I1 у славян) имеют снип М253, все “южные славяне” (I2a у славян) имеют снип L460, все “южно-балтийские славяне” и “угро-финны” — снип М46 (Tat), хотя многие южно-балтийские славяне имеют еще одну мутацию, L550, которой нет у финно-угорской ветви.
Каждый из перечисленных родов имеет свою структуру, свои ветви популяций, каждая из ветвей имеет свою историю. Например, в Европе насчитывается 38 ветвей гаплогруппы R1a, различающиеся по характерной картине мутаций в гаплотипах.
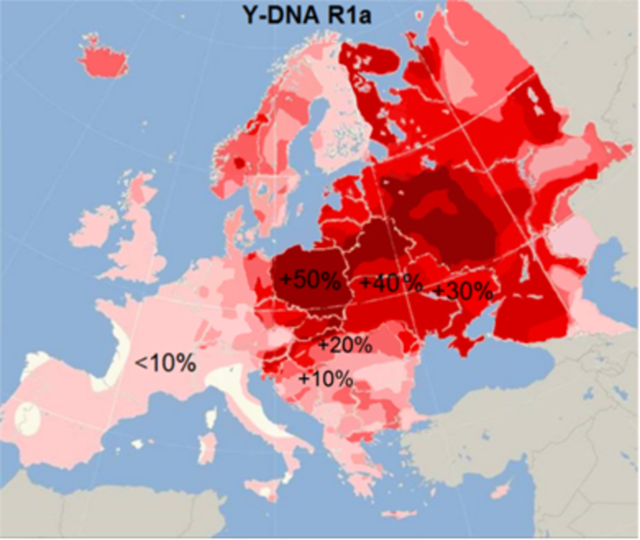
Распространение гаплогруппы R1a Материал из Википедии — свободной энциклопедии
Другая характерная гаплогруппа у славян — I, причем в двух подгруппах, I1 и I2, представленных среди славян неодинаково. Обе подгруппы распространены по всей Европе, причем I1 больше на севере Европы, I2 — больше на юге, в особенности на Балканах, где она часто является наиболее представительной по численности гаплогруппой, потому ее наиболее часто встречающийся там субклад (почти 100% от балканских гаплотипов группы I2) получил название «динарского» (индекс I2a1b) . Этот же субклад почти исключительно представлен у всех славян Восточной Европы. Это, наряду с I1 — древнейшая гаплогруппа Европы. Сама же гаплогруппа I появилась (путем соответствующей мутации М170) примерно 40 тысяч лет назад, и примерно 30 тысяч лет назад она разошлась на подгруппы I1 и I2.
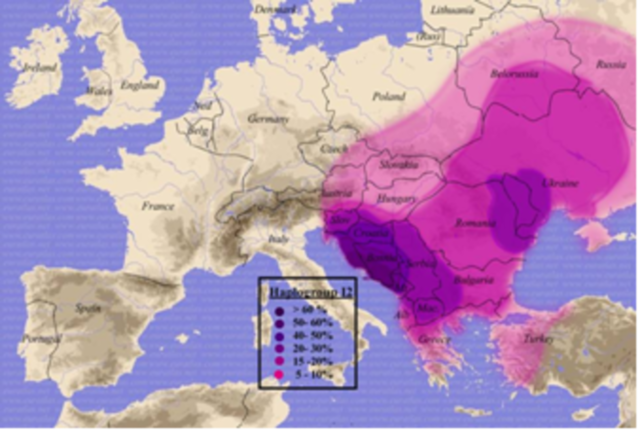
Распространение гаплогруппы I2 Материал из Википедии — свободной энциклопедии
Наконец, еще одна из наиболее представленных у славян гаплогрупп, N1c1, встречается в европейской части России в среднем в 14% случаев, но от Пскова и выше ее уже более 30%. Соответственно, на юге и на западе России этой гаплогруппы относительно мало. Она появилась у славян последней, в начале-середине первого тысячелетия нашей эры, пройдя длинный миграционный путь с Урала, а туда — со стороны Алтая через Сибирь. Заняло это по меньшей мере 10 тысяч лет, а сама гаплогруппа N разошлась с гаплогруппой О из сводной (родительской) гаплогруппы NO-M214 примерно 20 тысяч лет назад, когда у одного из носителей NO в Y-хромосоме произошла мутация М231, которая и положила начало 20-тысячелетней истории рода N.
Носители гаплогруппы R1a примерно 3500 лет назад прибыли в Индию как арии. На это есть целый ряд указаний. Во-первых, направления миграционных потоков гаплогруппы R1a, идущих из Европы через всю Русскую равнину в Индию; во-вторых, датировки этого миграционного пути, которые занимают промежуток времени от 4600 лет до 3500 лет назад, то есть от середины III до середины II тыс. до н. э.; в третьих, тот факт, что до 72% высших каст Индии занимают именно носители гаплогруппы R1a. Ни гаплогруппы I, ни N в Индии нет, как нет и гаплогруппы R1b, которая в настоящее время составляет примерно 60% Центральной и Западной Европы. В то же время гаплогруппа R1a составляет в среднем 48% этнических русских, доходя до 63% в южных областях России (Орловская, Курская, Белгородская области)».
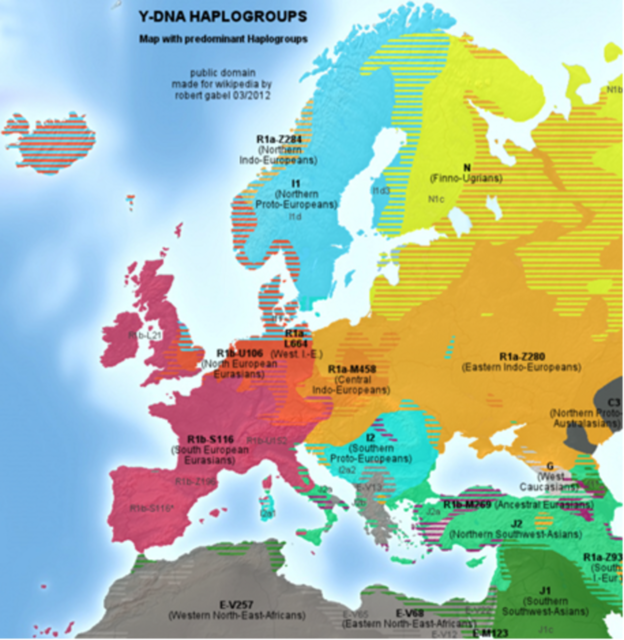
Расспространение гаплогрупп В Европе, Северной Африке и на Ближнем Востоке. Карта Роберта Гэйбла[4]
Важное примечание к карте Анатолия Клёсова: "Как следует из надписи в ее верхней части, эта карта представляет только «доминирующие» гаплогруппы. Поэтому на Балканах, например, гаплогруппа R1a на карте вообще не представлена, хотя ее доля составляет в Сербии 33%, Хорватии 24%, Боснии-Герцеговине 55%[5]. Далее, языковые группы на карте обозначены во многих случаях неверно — так, балты (литовцы, латыши) не являются «финно-уграми», они говорят на языках индо-европейской группы; то, что носители гаплогруппы I1 являются «северными прото-индоевропейцами», является ошибкой — никому не известно, на каких языках говорили скандинавы во времена прото-индоевропейских языков; это же относится и к гаплогруппе I2, которых автор карты обозначил как «южных прото-индоевропейцев». Это сейчас носители гаплогруппы I2a на Балканах и сопряженных территориях говорят на языках индоевропейской группы, как и скандинавы гаплогруппы I1, но назвать их «прото-индоевропейцами» никак нельзя. Клёсов делает типичную для популяционной генетики ошибку — приравнивает то, что наблюдается сейчас, с тем, что было (но что мы не знаем) многие тысячелетия назад. Это — еще одна причина, почему популяционная генетика — это не ДНК-генеалогия, у нее не тот методологический аппарат". (комментарий Анатолия Клёсова)
Согласно Клёсову сейчас существует две превалирующие по численности гаплогруппы у славян Балкан и Восточной Европы — I2а и R1a, и все гаплотипы славян гаплогруппы I2a (на примере сербов, македонцев, черногорцев, боснийцев, хорватов, словенцев, русских, украинцев, белорусов, словаков, чехов, а также поляков, греков, болгар, литовцев, латышей) — все принадлежат одной четкой ветви, под названием «динарская» (I2a1b по современной номенклатуре), являющейся на самом деле восточно-европейской ветвью, которая в свою очередь расходится на ветви. Их возраст примерно 2200 лет, но эти восточно-европейские ветви составляют всего треть от всех гаплотипов I2а в международных базах данных. Как пишет Анатолий Клёсов, напротив нее на дереве гаплотипов находится другая ветвь гаплогруппы I2а, почти такого же размера (численности) , чья родина почти исключительно Британские острова и чей возраст около 5000 лет. Британские острова являются также родиной последней трети гаплотипов группы I2а.
Согласно Клёсову, датировка британских гаплотипов примерно совпадает с прибытием в Европу эрбинов, носителей гаплогруппы R1b, началом эпохи колоколовидных кубков (4800 лет назад) с их стремительным — по историческим меркам — заселением Европы между 4800 и 4000 лет назад, и потому вероятным представляется истребление эрбинами автохтонных европейских родов-гаплогрупп, в результате чего I2а переселялись на запад, на Острова и на восток, на Балканы (прохождение бутылочного горлышка 2350 лет назад), тогда как R1a переселялись на Русскую равнину (4800–4600 лет назад), а G2 через Малую Азию на Кавказ (4500 лет назад).
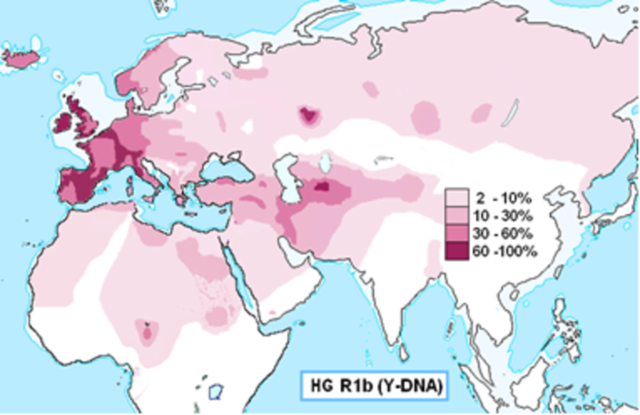
Распространение гаплогруппы R1b Материал из Википедии — свободной энциклопедии
Сейчас гаплогруппа G2 является наиболее многочисленной среди кавказского мужского населения северного и западного Кавказа (осетины, абхазы, черкесы, адыги).
На востоке славянский род I2а пошел в рост только в конце прошлой эры, то есть 2350 лет назад, и они сейчас и составляют большинство балканского населения. Так, согласно исследованиям Анатолия Клёсова, в настоящее время в составе сербского народа доминирующими гаплогруппами являются I2 (30–40%) и R1a (15–20%) , тогда как более минорными (менее 10% от общего состава) являются другие гаплогруппы как J2, G2, E1, I1a.
В настоящее время особенно высокий процент концентрации гаплогруппы I2а наблюдается в Боснии-Герцеговине, в южной Далмации в Хорватии, а также в Сербии и настолько высокая концентрация этой гаплогруппы наблюдается в Европе лишь в районе Молдовы и Румынии, и относительно высокий процент этой гаплогруппы обнаружен на Украине и в Белоруссии, 21% и 18%, соответственно[6].
Важную роль в истории славян сыграла гаплогруппа R1a, чьи ветви имеют «карпатский след», и ее северо-карпатская ветвь (в Сербии) датируется 2150±300 лет назад (сама ветвь состоит из двух подветвей, которые образовались примерно 2400 лет назад), балто-карпатская ветвь (Словения и Хорватия) 2200±250 лет назад (сама ветвь образовалась 4300 лет назад), а восточно-карпатская (Словения и Хорватия) 2400±300 лет назад, она тогда же и образовалась.
Можно еще раз процитировать Клёсова: «Носители гаплогруппы R1a приобрели славу как легендарные арии — в Малой Азии, на Ближнем Востоке, в Иране, в Индии. Это они, как показывает анализ данных ДНК-генеалогии в содружестве с археологией, изобрели боевые колесницы, воспетые в гимнах Индии — Ригведе, Рамаяне. Носители гаплогруппы G2 ушли через Малую Азию в Месопотамию, Иран, на Кавказ, и сейчас на западном Кавказе они составляют большинство популяций у осетин, абхазов, черкесов, адыгов/шапсугов».
Разница между историей гаплогрупп I2а и R1a, согласно Клёсову, заключалась в том, что носители I2а на Балканах и на Русской равнине древнее 2350 лет назад полностью пропали (во всяком случае, пока не обнаружены), тогда как носители R1a прослеживаются до глубин 5 тысяч лет назад.
Глубже по времени идет генеалогический обрыв, который восстанавливается только при перекрестном анализе ветвей и достигает как минимум 7–9 тысяч лет. Тем самым, основную историю славян, начиная от 4500 лет назад, создавал род R1a.
Можно предположить, что в формировании сербского этноса важную роль сыграли и другие гаплогруппы, что конечно нуждается в дальнейших исследованиях, так же как нуждается в исследовании и происхождение любого иного этноса. По состоянию исследований на сегодняшний день, в Сербии доминирует гаплогруппа I2a (33%), за ней следуют E1b (18%) и R1a (16%), далее I1, J2 и R1b (по 8%), в сумме это уже более 90%, остальные гаплогруппы — минорные по численности (I2b, G2a, J1, Q, T, N1c1).
Практически все современные народы различаются по генотипу (набору генов), по фенотипу (физическому отражению этого набора генов) и по гаплогруппам (отражение принадлежности к роду), и представляют собой произведение смешения различных рас и этносов.
Так, испанцы — продукт смешения как минимум трех народов — иберов, кельтов и германцев, и согласно профессору Анатолию Клёсову, нынешние испанцы на две трети имеют гаплогруппу R1b (среди басков — 85%), которая прибыла в результате длинных тысячелетних миграций со стороны Центральной Азии и далее Ближнего Востока, но является европеоидной по происхождению гаплогруппой. Следует учесть, что само понятие «европеоиды» не имеет отношения к территории, это — определение комплекса антропологических характеристик, которые в настоящее время больше всего выражены в Европе. Проще тут говорить об арийском облике, тем более что ныне существует предположение что «европеоиды» или проще говоря арийцы своей Родиной имели Сибирь и в Европу попали в ходе миграций из Сибири.
Учитывая современные исследования ДНК генеалогии можно предположить, что процент гаплогрупп у нынешних сербов и условно говоря «протосербов» мог значительно различаться, как и у прочих современных народов и их условных предков — «протонародов».
Однако на данный момент тяжело определить, какую гаплогруппу имели протосербы, а какую — представители иных народов, населявших Балканы, тем более, что гаплогруппа жителей древних цивилизаций Лепеньского Вира, Старчево и Винчи также пока с определенностью не известна, также как неизвестно были ли они теми «протосербами».
На Балканах живут представители различных рас в том понимании, которое было привычно для XIX — начало XX веков, когда под расами понимались антропологические типы изучавшиеся этнической антропологией, что ныне нередко осуждается (как правило, без оснований) как расизм. Вместе с тем наряду с изучением антропологических особенностей тогдашняя рассология изучала и психологические особенности вышеупомянутых типов или рас. Данные психологические особенности могли достаточно сильно различаться, точно также как и антропологический облик различных типов, даже внутри одного народа.
В Сербии в 1914 году вышла работа «Единство и психические типы динарских южных славян» известного историка и географа Йована Цвиича[7] в которой Цвиич употребляет понятие этнопсихологических профилей, дабы охарактеризовать психологические особенности, присущие тем или иным общностям южных славян. Под последними Цвиич понимал в своей работе прежде всего хорватов и сербов, включая сербов исламской вероисповести.
Сам Цвиич считал что среди «югославов» существует три главных этнопсихологических типа-тип «горно-динарский», разделявшийся Цвиичем на два подтипа: «северный» и «южный», тип «приморско-динарский» и тип «панонский» с тем что границы их преимущественного распространения не совпадали с этническими границами[8].
Эти типы Цвиич дополнительно разделял на различные группы составляя достаточно сложную мозаичную картину состояния тогдашних «югославов».
Вместе с тем следует учитывать, что и доныне еще можно обнаружить антропологические различия различных региональных типов даже в физическом облике — форме черепов, в росте, в телосложении, в цвете и форме волос (прямые или кучерявые). В данном случае Балканы, и в первую очередь сербы с их до сих пор сильными родственными связями, представляют собою достаточно широкое поле для исследований.
В бывшей Югославии сохранились различные антропологические типы славян, и так же, как у русских, в сербском народе существовало несколько антропологических типов, сильнее различавшихся между собою в отличие от русских.
Особенно это характерно для различных горных районов, и пресловутое смешение с турками никакого отношения к этому не имеет, ибо согласно исследованиям Вука Караджича в той же Шумадии в Сербии в начале XIX века большинство населения было либо переселенцами, либо потомками во втором-третьем поколении переселенцев из Черногории, где смешение при турках было минимальным.
Данные различия достаточно очевидны при мало-мальски серьезном исследовании таких районов с установлением условного общего типа.
В конце концов соседние сербам болгары, также относящиеся к южным славянам, по мнению Йована Цвиича в среднем куда сильнее отличались от «югославян» в том числе как внешним обликом, так и психологическим профилем, что, собственно, достаточно очевидный факт.
Также широко известен исторический факт, что после эпидемии чумы в VXII–XVIII веках в Романии и всей соседней области вокруг Сараево фактически был заново заселен переселенцами из Черногории, и оттуда волна переселенцев позднее пошла на запад и север Боснии и Герцеговины.
Различия во внешнем физическом облике в той же Боснии и Герцеговине сохранились до сих пор, и герцеговинцы в общем отличаются ростом, а нередко и формой черепов от боснийцев.
В данном случае необходима программа по проведению исследования гаплогрупп Балкан, с параллельным исследованием историй происхождения родов самих исследуемых особей. Конечно, подобное исследование требует и исследования могильников древних цивилизаций, существовавших на Балканах, с выяснением культуры и языка этих цивилизаций.
Все же можно предположить, что со временем какая-то гаплогруппа, являвшаяся основной для того или иного народа, стала замещаться в силу тех или иных причин, как, например, более высокой рождаемости представителей иной гаплогруппы, либо общественных потрясений. Примеры, когда в состав одной этнической группы вливались какие то другие этнические группы, известны и по истории Сербии XIX века, когда в состав сербского народа вошло большое число влахов и цинцаров, отличавшихся от сербов как культурой и языком, так и нередко физическими данными.
Тут интересно различие антропологического типа, или, проще говоря, среднего физического облика сербов с горных массивов Загорья (Калиновик) и Озрена (Петрово село), отделенных друг от друга всего двумя сотнями километров. Оба типа отличаются от сербов, допустим, Южной Сербии (Прокуплье, Куршумлия, Вранье) повышенным числом светловолосых особей, с тем, что в первом случае имеют место особи высокого роста и прямых волос, а во втором случае особи более низкого в среднем роста и повышенным процентом особей кучерявых волос.
В данном случае речь идет не об индивидуальных различиях а о различии между общими антропологическими типами этих двух общностей, с тем, что подобные различия в большей мере характерны как раз для сельских жителей, поколениями жившими на этих просторах.
Имеют они общее происхождение или все таки различное на данный момент оценить невозможно, ибо в бывшей Югославии исследований геномов населения просто не проводилось в широких масштабах.
Вышеприведенные особенности антропологических типов, как и этнопсихических профилей, дают основание предположить о том, что на Балканах произошло слияние нескольких рас в их как уже упоминалось «узком» понимании, и вполне возможно, что сербы, сохранив язык и имя оригинальных «протосербов», в то же время ассимилировали ряд других рас или хотя бы одной расы.
В конце концов, именно это и произошло с древними «ариями» в Индии, которые испытали такое же влияние, хотя язык древних «ариев» санскрит в Индии сохранился. Впрочем, многие потомки древних ариев, особенно в Северной Индии, практически полностью сохранили свою «европеоидность», включая и светлый цвет кожи.
Тем самым вполне возможным представляется то, что в отдельных сербских районах мог в большем проценте сохраниться тип протосербов, или, условно говоря, протославян, которые в древности в значительной мере отличались и физическими и психическими особенностями от нынешних сербов, а также от современных русских, тоже переживших процессы ассимиляций иных рас. Хроники былой Ромейской империи, хорошо знакомой со славянами, дают картину славян во многом иного физического облика и психического склада, нежели то, что привычно ныне, и, возможно, причины тому заключаются не только в культуре и воспитании.
Ничего удивительного в этом нет, так как само возникновение многих народов в древности происходило в ходе смешения различных племен и каждое племя вносило свои психические и антропологические особенности. К тому же существуют примеры, когда те или иные народы усваивали чужие языки и культуры, тогда как возможно (но не доказано) сохраняли свои этнопсихологические профили.
В данном случае современная ДНК-генеалогия, если объединит свои усилия с археологией, то, возможно, определит судьбу ныне уже исчезнувших народов либо их происхождение, а может предоставить и более широкие возможности для выяснения происхождения многих современных народов, тем более что возможно, что иные из них, сохранив язык и имя, не смогли сохранить гены древних протонародов .
В данном случае следует учитывать возможность того, что в истории известны случаи, когда тот или иной этнос ассимилировался другим этносом, несмотря на свой более высокий культурный уровень. Например, норманы усвоили во Франции французский язык, который использовали несколько веков и в Англии, после ее завоевания, а затем полностью влились в английский этнос.
О судьбе исчезнувших народов сегодня строят догадки немногие исследователи и писатели. Македонский писатель Панче Хаджи-Андонов в своей книге «Потерянные племена Израиля»[9] сделал предположение что скифы как раз и были потомками одного из пропавших колен Древнего Израиля, и по его мнению, современные русские являются потомками полуколена Ефрема. Конечно, такие заключения как и другие, опирающиеся на Библию, сегодня принято считать антинаучными, однако когда-то и существование Ниневии многими учеными XIX века воспринималось как фантастика из библейских текстов, пока она не была обнаружена в холме Куюнджик в 1849 году археологом Остином Генри Лэйярдом. Предположение Панче Хаджи-Антонова кажется совершенно безосновательным, потому что современные русские имеют совершенно иные ДНК-линии (гаплогруппы и их подгруппы, называемые субкладами), чем современные евреи. Однако неизвестно какие гаплогруппы-субклады имели пропавшие «колена израилевы» и потому неизвестно в каких народах можно обнаружить их потомков. Очевидно что среди современных евреев их нет иначе бы эти колена не назвались «пропавшими». В данном случае достаточно хорошо известно как от историков церкви так и от иудейских историков, что в год земной жизни Иисуса Христа древние иудеи (евреями их начали назвать в Просвященном веке, а до этого они звались иудеи или juden по немецки) были в своей массе потомками колен Иуды и Венеамина, в небольшой мере и Левия. Хотя среди них уже тогда находился значительный процент ассимилированных потомков окружающих их народов, например, идумеян. Самым известным таким иудеем идумейского происхождения был царь Ирод. Так как иудаизм был не просто религиозным заветом но и заветом крови, относившейся исключительно к потомкам первых двенадцати колен Израилевых, то уже тогда существование ассимилированных потомков инородцев вызывало протесты в иудейской среде, в частности против того же царя Ирода.
Судьба «пропавших колен Израиля» традиционно привлекает к себе внимание историков. По сообщению русскоязычного журнала «Канон» выходящего в США в городе Сиэтл[10], в Израиле иерусалимский раввин Элиягу Авихиль издал книгу «Колена Израиля», в которой установил ареал распространения «потомков пропавших колен» в восточной Азии на территории современных Пакистана и Афганистана. Впрочем, этот вопрос был подробно рассмотрен в недавней книге А.А. Клёсова и Х.С. Саидова «Евреи и пуштуны Афганистана»[11], в которой доказывалось что древняя легенда о происхождении пуштунов от «пропавших колен израилевых» не имеет никаких оснований.
Современная ДНК-генеалогия утверждает что современные евреи, имеют несколько гаплогрупп, с тем что пока «наиболее еврейской» по численности считается гаплогруппа J1 которая обнаружена и у арабов, однако наличие различных гаплогруп у евреев дает поле для различных теорий, тем более что никто не знает, какие гаплогруппы были у мифических «пропавших» десяти колен Древнего Израиля, уведенных в плен ассирийским царем Саргоном Вторым в 722–720 годах до н.э.
В недавней книге А.А. Клёсова «Кому мешает ДНК-генеалогия»[12] методами ДНК-генеалогии показано существование общего предка евреев и арабов гаплогруппы J1, который жил примерно 4000 лет назад.
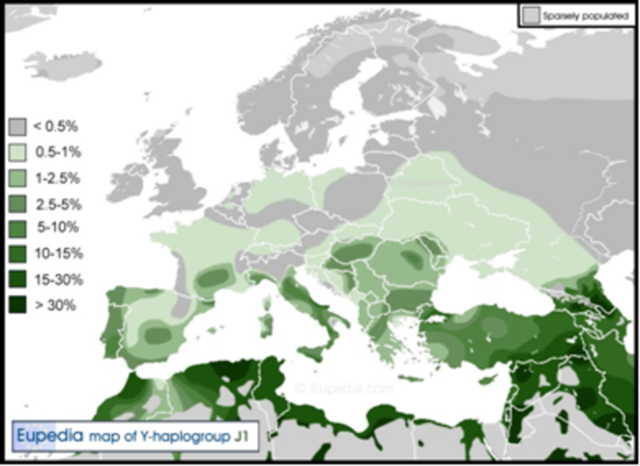
Распространение гаплогруппы J1 Источник сайт Eupedia https://www.eupedia.com/
Там же показано, что общий предок евреев и арабов есть и в гаплогруппе J2, и в гаплогруппе R1a, и во всех трех гаплогруппа он жил примерно 4000 лет назад, однако наличие трех общих предков, да еще с несоответствующим Библии временным промежуткам противоречит учению Библии об Аврааме, что принуждает либо отказываться от Библии, либо продолжать исследования.
В среде современных евреев относительно много представителей других гаплогрупп, и по самой крупной выборке из работы Бехара[13] у евреев-ашкенази было найдено 10% гаплогруппы R1b, 8% гаплогруппы R1a, 6% гаплогруппы Q1b, 19% гаплогруппы J1, 8% гаплогруппы G2, 17% гаплогруппы E1b1b1 и 19% гаплогруппы J2[14]. И хотя за последние 12 лет эти оценки несколько изменились, но не столь существенно, чтобы этому уделять особое внимание.
Помимо этого, неясна количественная роль в формировании еврейского народа гаплогруппы J2 , ибо у иудейских «коэнов», предположительно потомков Аарона, брата Моисея и потомка Афраама, согласно Библии, были обнаружены как гаплогруппа J-1 так и гаплогруппа J-2, а также еще десяток относительно минорных гаплогрупп[15]. Возникает вопрос о том какая же из этих гаплогрупп является «исконной» для Древнего Израиля.
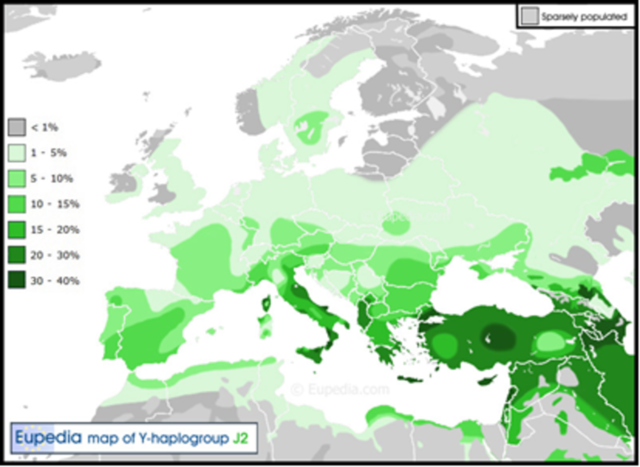
Распространение гаплогруппы J-2. Источник сайт Eupedia https://www.eupedia.com/
Сам Клёсов, располагавший большой базой данных ДНК «коэнов», которые среди всех евреев самые щепетильные в ведении родословной, обнаружил, как уже упоминалось, немалое количество ДНК-линий коэнов, помимо J1 и J2, также R1a, R1b, E1b, G1, G2a, H1, L, Q, R2a, T1a и несколько других[16]. На основе данных из работы Бехара[17] из двух ветвей «коэнов», одна из гаплогруппы J1, появилась примерно 1000 лет назад, тогда как другая ветвь «коэнов» из гаплогруппы J2 появилась три с половиной тысяч лет назад[18]. Правда расчеты тогда велись без достаточного знания скоростей мутаций, поэтому получающиеся датировки значительно искажены.
При этом имелось еще несколько подвидов «коэнов» гаплогруппы J2, и на основании работы Хаммера (Hummer. et al. 2009) [19] профессор Клёсов определил, что один из трех подвидов «коэнов» гаплогруппы J2, носящий обозначение J-2a-M410 появился примерно полторы тысячи лет назад, тогда как подвид J-2b-M12 имел возраст 3150 (±290), с тем что до общего предка J2a-M410 и J2b-M12 было семь тысяч лет[20]. Данное обстоятельство находится в полном противоречии с Библией, ибо невозможно, чтобы одна большая часть потомков Аарона имела одного предка, а другая — другого, причем нелогично, что одни «коэны», как и основная масса современных евреев гаплогруппы J1, ближе к арабам, нежели к своим соплеменникам — «коэнам» из J2.
В данном случае невозможно воспринимать Библию «символически», ибо логичнее тогда ее вообще не воспринимать как исторический документ, хотя в таком случае теряется ее ценность и как Откровения.
Сама гаплогруппа J2, которая была предметом исследований, проведенных профессорами Гарвардского университета Пьером Залоа (Pierre Zalloua) и Спенсером Уэллсом (Spencer Wells) в рамках проекта “National Geographic Genographic Project” в 2004 году, была предметом и последующих исследований, не относящихся к евреям. Эти исследователи назвали данную гаплогруппу «финикийской», и, как пишет Ди Джиакомо[21] эта группа может охарактеризоваться как «средиземноморская» но не как «семитская».
Данная гаплогруппа широко представленная у греков (в первую очередь Кипра и Крита), у итальянцев (в первую очередь Сицилии), у испанцев, у мальтийцев, а также у турок (прежде всего в районе Анадолии), на Балканах-среди валахов, болгар и албанцев, на Кавказе в первую очередь среди чеченцев и ингушей (причем интересно что вайнахи-чеченцы и ингуши единственный народ у которого данная древняя гаплогруппа доминирует), а также у осетин, черкесов, абхазцев, балкарцев, азербайджанцев, грузин, кумыков, армян, лазов, мегрелов, а также в Иране у персов, у арабов (Палестины, Ирака, Сирии, Омана, Саудовской Аравии, Иордании, Ливана), у хазарейцев в Афганистане и у калашей в Нуристане, как и у некоторых каст в Индии как и у индийских мусульман-шиитов и у народности саамы в Скандинавии с тем что ее можно найти и у многих других народов, в том числе среди современных сербов, украинцев и русских. Сама ДНК генеалогия в России находиться пока в зачаточном состоянии и пока еще нельзя делать какие-либо выводы о происхождении тех или иных народов. Тем более не сделаны выборки у отдельных небольших этнических группы как например у донских и кубанских казаков, у кержаков в Сибири,у таких малоизвестных народах Кавказа как табасаранцы ,у мелеузских татар, у селькупов наконец у племен гуцулов, бойков и лемков на Украине. Поэтому нельзя сделать уже какие то общие выводы по движениям представителей тех или иных гаплогрупп по территории Восточной Европы, Сибири и Средней Азии с целью выяснения очагов распространения предков современных народов,в том числе и русского.
Источники
- Сайт “ Family Tree DNA ” http://www. familytreedna. com/
- Материалы международной научной конференции «У источника культуры и науки». Белград. 2012 год
- Proceedings of the Academy of DNA Genealogy) » (Volume 5, No. 10 October 2012
- «Јединство и психички типови динарских јужних словена». Јован Цвијић. ”Слободна књига-издања Владимира Максимовића”. Београд. 1999 г.) .
- “Izgubljena plemena Izraela”. Hadži-Andonov. ”Pešić i sinovi”. Beograd. 2006 g.
- «Судьба колен израилевых». Иосиф Каменецкий. «Канон». Номер за сентябрь от 2005 года
- «Contrasting patterns of Y chromosome variation in Ashkenazi Jewish and host non-Jewish European populations». Doron M Behar; Daniel Garrigan; Matthew E Kaplan; Zahra Mobasher; Dror Rosengarten; Tatiana M Karafet; Lluis Quintana-Murci; Harry Ostrer; Karl Skorecki; Michael F Hammer. Human genetics 2004.
- «Происхождение человека». Анатолий Клёсов, Андрей Тюняев. «Белые Альвы». Москва. 2010 г.
- «Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood». Hammer MF, Behar DM, Karafet TM. Human Genetics . (November 2009) .
- Klyosov, A.A. (2009) A comment on the paper: Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood by M.F. Hammer, D.M. Behar, T.M. Karafet, F.L. Mendez, B. Hallmark, T. Erez, L.A. Zhivotovsky, S. Rosset, K. Skorecki, Hum. Genet. 126, No. 5, 719-724).
- Клёсов, А.А. «ДНК-генеалогия от А до Т». М., Книжный мир, 2016, 475 с.
- Клёсов, А.А. «Кому мешает ДНК-генеалогия», М., Концептуал, 2016, 848 стр.
- Клёсов, А.А., Пензев, К.А. «Арийские народы на просторах Евразии». М., Книжный мир, 2015, 3
Примечания
[1] Сайт Family Tree DNA // www. familytreedna.com
[2] Материалы международной научной конференции «У источника культуры и науки». Белград, 2012 год
[3] Proceedings of the Academy of DNA Genealogy. Vol. 5. 2012. № 10.
[5] www.eupedia.com/europe/european_y-dna_haplogroups.shtml.
[6] www.eupedia.com/europe/european_y-dna_haplogroups.shtml.
[7] Јован Цвијић. Јединство и психички типови динарских јужних словена. Београд: Слободна књига-издања Владимира Максимовића, 1999.
[8] Јован Цвијић. Јединство и психички типови динарских јужних словена. Београд: Слободна књига-издања Владимира Максимовића, 1999.
[9] Hadži-Andonov. Izgubljena plemena Izraela. Beograd: Pešić i sinovi, 2006.
[10] Иосиф Каменецкий. Судьба колен израилевых // Канон. 2005. Сентябрь.
[11] Клёсова А.А., Саидова Х.С. Евреи и пуштуны Афганистана. М.: Концептуал, 460.
[12] Клёсов А.А. Кому мешает ДНК-генеалогия. М.: Концептуал, 2016. 848 с.
[13] Doron M Behar; Daniel Garrigan; Matthew E Kaplan; Zahra Mobasher; Dror Rosengarten; Tatiana M Karafet; Lluis Quintana-Murci; Harry Ostrer; Karl Skorecki; Michael F Hammer. Contrasting patterns of Y chromosome variation in Ashkenazi Jewish and host non-Jewish European populations // Human genetics. 2004.
[14] Doron M Behar; Daniel Garrigan; Matthew E Kaplan; Zahra Mobasher; Dror Rosengarten; Tatiana M Karafet; Lluis Quintana-Murci; Harry Ostrer; Karl Skorecki; Michael F Hammer. Contrasting patterns of Y chromosome variation in Ashkenazi Jewish and host non-Jewish European populations // Human genetics. 2004.
[15] Клёсов А.А. ДНК-генеалогия от А до Т. М.: Книжный мир, 2016. С. 209–222.
[16] Клёсов А.А. ДНК-генеалогия от А до Т. М.: Книжный мир, 2016. С. 210–213.
[17] Doron M Behar; Daniel Garrigan; Matthew E Kaplan; Zahra Mobasher; Dror Rosengarten; Tatiana M Karafet; Lluis Quintana-Murci; Harry Ostrer; Karl Skorecki; Michael F Hammer. Contrasting patterns of Y chromosome variation in Ashkenazi Jewish and host non-Jewish European populations // Human genetics. 2004.
[18] Doron M Behar; Daniel Garrigan; Matthew E Kaplan; Zahra Mobasher; Dror Rosengarten; Tatiana M Karafet; Lluis Quintana-Murci; Harry Ostrer; Karl Skorecki; Michael F Hammer. Contrasting patterns of Y chromosome variation in Ashkenazi Jewish and host non-Jewish European populations // Human genetics. 2004.
[19] Hammer MF, Behar DM, Karafet TM. Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood // Human Genetics. 2009. November.
[20] Klyosov A.A. A comment on the paper: Extended Y chromosome haplotypes resolve multiple and unique lineages of the Jewish priesthood by M.F. Hammer, D.M. Behar, T.M. Karafet, F.L. Mendez, B. Hallmark, T. Erez, L.A. Zhivotovsky, S. Rosset, K. Skorecki // Human Genetics. 2009. 126. № 5. P. 719–724.
[21] Di Giacomo. Y chromosomal haplogroup J as a signature of the post-neolithic colonization of Europe // Hum Genet. 2004. October.)















2 комментария